L’appareil reproducteur des angiospermes (qui sont des spermaphytes) est la fleur qui après fécondation donne naissance à la graine et au fruit.
Organisation générale de la fleur
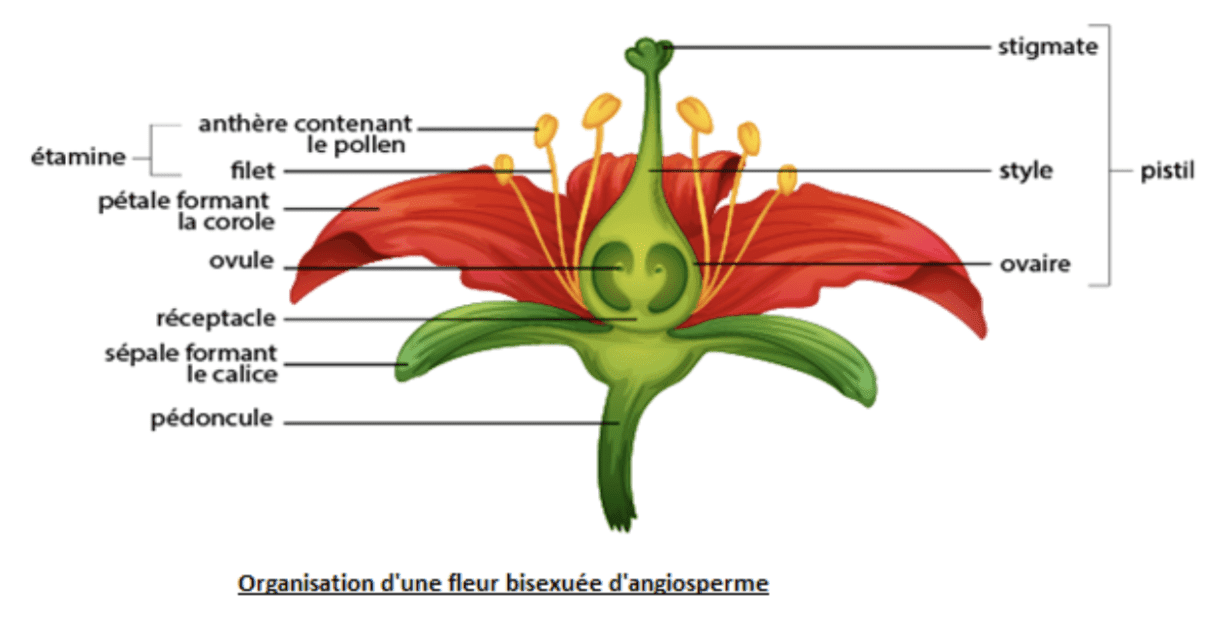
La fleur est portée par un axe dressé, le pédoncule, terminé par un renflement : le réceptacle qui porte les pièces florales. Ces pièces florales peuvent être stériles ou fertiles.
Les pièces stériles
L’ensemble des pièces protectrices stériles d’une fleur forment le périanthe. Il comprend : le calice (ensemble des sépales, généralement de couleur verte) et la corolle (ensemble des pétales, de couleurs variables).
Les pièces fertiles
Des parties mâles (l’androcée) et des parties femelles (le gynécée) :
L’androcée
C’est l’ensemble des étamines. Chaque étamine comprend un filet étroit et une anthère. Cette dernière est formée de plusieurs sacs polliniques ou sporanges séparés par des fentes de déhiscence dont l’ouverture permet la libération des grains de pollen contenus dans l’anthère à la maturité.
Le gynécée ou pistil
Il comprend un ovaire (formé de carpelles) renfermant des ovules reliés à un placenta, un style terminé par un stigmate. Chaque ovule contient un sac embryonnaire à maturité.
NB : Les fleurs possédant des organes mâles et femelles sont dites hermaphrodites ou bisexuées ou dioïques, alors que celles qui ne possèdent qu’un seul type d’organes sont dites unisexuées ou monoïques (ex : le rônier, le dattier…).
Les organes reproducteurs mâles : les étamines
Structure des anthères et des grains de pollen
L’anthère présente un parenchyme, des faisceaux conducteurs et 4 sacs polliniques ou sporanges entourés de l’extérieur vers l’intérieur d’un épiderme, d’une assise mécanique et d’une assise nourricière. Dans les sacs polliniques se trouvent des grains de pollen constitués de deux cellules, une cellule reproductrice ou génératrice (de petite taille et à noyau allongé) et une cellule végétative (de grande taille et à noyau arrondi). Le grain de pollen est délimité par deux membranes : l’exine (membrane externe dure, ornementée, avec des pores et parfois des épines) et l’intine (membrane interne mince et souple).
Formation des grains de pollen
Les cellules mères des grains de pollen, diploïdes (2n chromosomes), subissent une méiose et donnent 4 microspores haploïdes (n chromosomes). Ces microspores sont regroupées par la paroi de la cellule mère et forment des tétrades (4 microspores). Chaque microspore élabore son exine et son intine. Le noyau se divise pour donner la cellule reproductrice et la cellule végétative (la cellule reproductrice sera par la suite incluse dans la cellule végétale). La paroi qui unissait les tétrades s’hydrolyse et libère dans le sac pollinique les microspores devenus grains de pollen.
Structure du gynécée ou pistil
Le pistil est constitué du stigmate qui est relié à l’ovaire par le style. L’ovaire est formé d’un ou de plusieurs carpelles soudés (3 carpelles chez le Lis) contenant les ovules. Chaque ovule est relié au carpelle par un placenta sur lequel se fixe le funicule. L’intérieur de l’ovule présente un tissu, le nucelle qui est recouvert d’un ou de deux téguments (primine et secondine). Sur le bout opposé à la chalaze, les téguments laissent un petit orifice, le micropyle. Dans le nucelle et vers le micropyle se trouve le sac embryonnaire. L’ovule peut présenter plusieurs formes : droit (orthotrope), courbé (campylotrope) et renversé (anatrope).
Formation du sac embryonnaire
Lors du développement du nucelle, une cellule se différencie par sa grosseur et son contenu, c’est la cellule mère du sac embryonnaire. Elle subit une méiose pour donner 4 macrospores ou mégaspores haploïdes. Les trois dégénèrent et celui qui survit augmente de taille et son noyau subit 3 mitoses successives pour donner 8 noyaux. Le cytoplasme produit ensuite une membrane qui délimite vers le pôle micropylaire une oosphère encadrée de deux synergides, vers le pôle chalazien 3 antipodes et au centre une cellule constituée de 2 noyaux, les noyaux polaires ou noyaux du sac embryonnaire (qui fusionneront pour donner un noyau diploïde, le noyau central).
La pollinisation, la germination du grain de pollen et la double fécondation
La pollinisation
C’est le transport passif du grain de pollen de l’anthère jusqu’au stigmate (qui le retient grâce à son aspect gluant et à ses papilles). Elle est directe quand le pollen provient de la même fleur ou croisée quand le pollen vient d’une autre fleur. Le transport du pollen est généralement assuré par le vent (anémogamie), les insectes (entomogamie), les oiseaux (ornithogamie) ou l’eau (hydrogamie).
Germination du grain de pollen et croissance du tube pollinique
Le grain de pollen posé sur le stigmate absorbe de l’eau par osmose et se gonfle. Sous l’effet de la pression due à la turgescence, l’intine et une partie du cytoplasme de la cellule végétale font saillie au niveau des pores de l’exine et donnent un tube pollinique, c’est le début de la germination.
Le stigmate émet une substance (acide borique) qui diffuse jusqu’aux grains de pollen (les grains éloignés du stigmate ne sont pas atteints). Cette substance oriente la croissance du tube pollinique vers le stigmate, c’est le chimiotropisme positif.
La double fécondation
Au début de la germination le noyau de la cellule végétale se place à l’extrémité du tube pollinique (il dégénérera à la fin de la croissance du tube pollinique), il sera suivi par la cellule génératrice qui subit une mitose pour donner 2 gamètes mâles (anthérozoïdes ou spermatozoïdes) plus ou moins spiralés. Quand le tube pollinique atteint le sac embryonnaire, un des spermatozoïdes fusionne avec l’oosphère pour donner l’œuf principal ou œuf embryon (diploïde, 2n), l’autre spermatozoïde fusionne avec le noyau de la cellule centrale pour donner l’œuf accessoire ou œuf albumen (triploïde, 3n). Donc il y a une double fécondation qui est caractéristique des spermaphytes.
Mécanismes de formation de la graine et du fruit
Formation de la graine
La transformation de l’ovule fécondé se fait en trois temps :
- L’œuf accessoire se divise plusieurs fois et rapidement pour donner un tissu de réserve, l’albumen qui s’agrandit en digérant le nucelle.
- L’œuf embryon se divise plusieurs fois pour donner l’embryon ou la plantule (radicule, tigelle et gemmule) et les cotylédons.
- Lorsque le fruit est mûr, le développement de la graine s’arrête, elle se déshydrate et entre en vie ralentie. Les téguments deviennent épais et résistants. Selon la localisation des substances de réserve, nous avons des graines sans albumen ou exalbuminées (réserves dans les cotylédons) et des graines à albumen (réserves dans l’albumen).
Formation du fruit
Les fruits ne sont produits que chez les angiospermes et sont issus du développement de l’ovule (qui donne la graine) et des parois de l’ovaire (qui donne le péricarpe chargé de substances sucrées chez les fruits charnus).
Chez les gymnospermes, il n’y a qu’une graine nue issue du développement de l’ovule.
Germination
Conditions de la germination
Pour germer la graine doit remplir des conditions internes (être saine, mûre, pas trop vieille) et des conditions externes favorables (besoin d’eau, de l’oxygène et une température idéale).
Manifestations de la germination
Manifestations morphologiques
La graine semée absorbe de l’eau et se gonfle. Son tégument se fend, la radicule se développe et s’enfonce dans le sol. La tigelle en particulier l’hypocotyle, s’allonge vers le haut entraînant les cotylédons, c’est « la levée » de la graine, qui est épigée chez le niébé et l’arachide, contrairement à la germination hypogée où l’hypocotyle ne s’allonge pas et les cotylédons restent dans le sol. Les cotylédons s’ouvrent, la seconde partie de la tigelle (l’épicotyle) se développe en même temps que les premières feuilles qui verdissent au contact de la lumière. Les cotylédons vont ensuite perdre leur volume, verdir, flétrir et tomber.
Manifestations physiologiques
Le gonflement de la graine entraine une absorption d’eau par le cytoplasme des cellules, la graine passe de l’état de vie ralentie à une vie active avec :
- la synthèse d’enzymes de digestion (amylase, lipases et de protéases) qui digèrent les réserves en des nutriments simples (glucose, acides gras et acides aminés) utilisables par la plantule ;
- une activité respiratoire intense se manifeste par un taux élevé de CO2 dégagé et une forte absorption d’O2, on note également une élévation de la température.
Ces deux activités permettent à la plantule de grandir à partir de ses réserves, elle est hétérotrophe à ce stade. La radicule se développe ensuite pour donner des racines capables d’absorber l’eau et les sels minéraux du sol, la gemmule donne des feuilles qui verdissent et la plantes devient capable de synthétiser ses propres nutriments par photosynthèse et devient autotrophe.
